Глава 11. Эволюция жизни
11.3. Механизмы видообразования
11.3.1. Генотип как система
Популяционная генетика – это раздел генетики, изучающий генетическое строение и динамику генетического состава популяций – группы организмов, принадлежащих к одному и тому же виду и занимающих ограниченную географическую область обитания. Именно этот раздел науки является основой синтетической теории эволюции, принятой в настоящее время большинством учёных. Генотип совместно с внешней средой определяет фенотип организма. Фенотипы, приспособленные к условиям места обитания, сохраняются в процессе естественного отбора; неприспособленные фенотипы подавляются. Для популяции в целом судьба отдельной особи значения не имеет.
 1
|
| Рисунок 11.3.1.1. Генетическое разнообразие популяций на примере цветков различной окраски |
Генофонд – это многообразие генов и аллелей, имеющихся в популяции. В каждой популяции, размножающейся половым путём, генофонд постоянно изменяется; популяция эволюционирует. Новые сочетания генов проходят непрерывный естественный отбор, определяющий в конечном итоге, какие гены будут переданы следующим поколениям.
Частотой аллеля называют отношение количества данных аллелей у всех особей к общему количеству аллелей в популяции. Частоту доминантного аллеля обычно обозначают буквой p, частоту рецессивного аллеля – буквой q. Если ген представлен двумя аллелями, то выполняется математическое равенство
Таким образом, зная частоту одного из аллелей, можно определить частоту и другого аллеля. Так, если частота доминантного аллеля равна 78 %, то частота рецессивного аллеля равняется q = 1 – p = 1 – 0,78 = 0,22 (или 22 %).
Для частот аллелей существует условие равновесия Харди–Вайнберга. Частоты доминантного и рецессивного аллелей остаются неизменными, если в популяции выполняются следующие условия:
- размеры популяции достаточно велики;
- спаривание и размножение особей происходит случайным образом;
- естественный отбор отсутствует (все генотипы одинаково приспособлены к внешним условиям);
- различные поколения не скрещиваются между собой;
- не возникает новых мутаций;
- отсутствует обмен генами с другими популяциями.
Невыполнение одного или нескольких из указанных условий может привести к изменению частоты аллелей и вызвать эволюционные изменения в данной популяции.
Используя символы p и q, можно по-новому записать схему моногибридного скрещивания.
 2
|
| Рисунок 11.3.1.2. Моногибридное скрещивание на языке частот |
 3
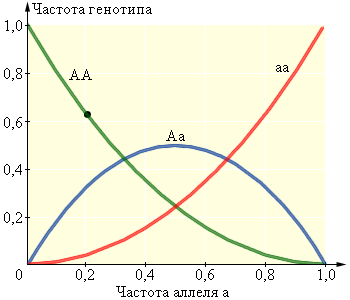
|
| Рисунок 11.3.1.3. Зависимость между частотами аллелей и генотипов |
Таким образом, при моногибридном скрещивании появляются три генотипа: AA с частотой p2 (гомозиготные особи с доминантным аллелем), Аа с частотой 2pq (гетерозиготные особи) и аа с частотой q2 (гомозиготные особи с рецессивным аллелем). Сумма частот аллелей равна единице:
Эта зависимость называется уравнением Харди–Вайнберга. Используя совместно это уравнение с уравнением p + q = 1, можно вычислить частоту, например, особей, гомозиготных по доминантному аллелю, зная количество носителей рецессивного фенотипа (то есть частоту особей, гомозиготных по рецессивному фенотипу). Пусть q2 = 0,0004. Тогда q = 0,02, p = 1 – q = 0,98, p2 = 0,9604, 2pq = 0,0392. Следствием уравнения Харди–Вайнберга является значительное превышение (часто на порядки) количества особей, в генотипе которых присутствует рецессивный аллель, над количеством особей с рецессивным фенотипом. Благодаря наличию значительного количества гетерозиготных особей в каждом поколении из генотипа исключается лишь малое количество рецессивных аллелей.
Многие рецессивные гены неблагоприятны для организма и исключаются из генотипа популяции. Другие, наоборот, полезны, благодаря чему частота гетерозиготного генотипа стабилизируется.
Как уже отмечалось, причиной изменений генофонда популяции являются половые рекомбинации генов (кроссинговер, независимое расхождение хромосом, случайное оплодотворение) и мутации. Половые рекомбинации не привносят в популяцию новые аллели; мутации вызывают появление новых аллелей и эволюционные изменения в популяциях. Существуют и другие причины генетической нестабильности, перечисленные ниже.
- Небольшая численность популяции может привести к дрейфу генов. Носителей рецессивного аллеля может оказаться настолько мало, что они исчезнут по причинам, не вызванным естественным отбором (например, по причине гибели в результате природного бедствия). Может получиться и обратная ситуация, когда частота рецессивного аллеля случайно вырастет. Это может никак не сказаться на популяции, привести к её вымиранию или наоборот стать благоприятным фактором.
- Структурные и поведенческие особенности, благоприятствующие какому-либо признаку (так, брачные игры у животных выигрывает обычно более крупный и сильный самец, а более интенсивно опыляются более яркие и крупные цветки), приводят к неслучайному скрещиванию, то есть к подавлению некоторых аллелей.
- Обмен генами между популяциями привносит, с одной стороны, новые гены в популяции, но, с другой стороны, уменьшает общее генетическое разнообразие видов и мешает эволюционным изменениям. Прерывание обмена генами между популяциями является важной предпосылкой для появления нового вида.

 |
|
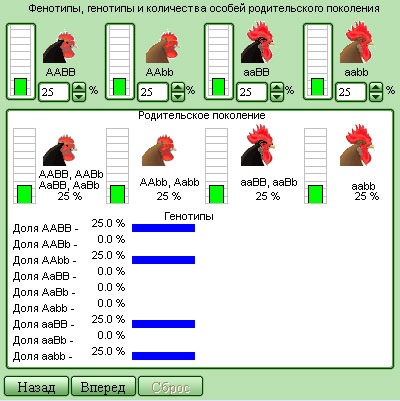
Модель 11.2.
Комбинативная изменчивость в популяции
|




